Activities carried out:
The WP1 was planned mainly as a field resampling of a large (ca. 144 ha) research site covered by six types of forest, which were established at the turn of the 1980s and 1990s by the team of researchers led by late prof. Janusz B. Faliński for the needs of the project CRYPTO (hereafter referred to as CRYPTO or survey 1990). The study plot was established in the strictly protected area of the Białowieża National Park, where forest ecosystems have not been influenced by direct human interference for at least the last 100 years. The resampling was precisely relocated owing to permanent markings of plot corners in the field (100mx100m) maintained since CRYPTO. All taxonomic groups of plants, most of lichens and many taxonomic groups of fungi were surveyed in the framework of the CRYPTO. The KlimaVeg project (hereafter also survey 2015) resampled the same 144 1-ha plots allowing for comparisons between the surveys at the scale of individual plots, plots grouped by the forest community and total study area. The KlimaVeg sampling limited its scope to lichens and plants (vascular plants, mosses, liverworts). Fungi were excluded from the resampling in the phase of preparation of the project because mycological studies require a different methodological approach. Vascular plants, bryophytes (mosses and liverworts) and lichens were resurveyed in all 144 plots and some general findings are presented below. The analyses of data obtained in the two additional resurveys carried out in the framework of the KlimaVeg project: phytosociological monitoring plots by Siemianówka water reservoir (northern part of the Białowieża Forest) and phenological monitoring plots are reportedin WP 4.
Results achieved:
Vascular plants. As expected, the site list of the vascular plants did not change considerably, the total species richness increased from 303 in 1990 to 320 species in 2015 with 24 species not confirmed in 2015 and 41 species recorded for the first time in 2015. The mean number of species per 1 ha plot increased from 73 ± 23 to 83 ± 23 species. Most of the species recorded as new in 2015 were characteristic for ruderal and segetal communities (e.g. Cirsium vulgare, Plantago major, Vicia hirsuta) and open grasslands (e.g. Holcus lanatus, Phleum pratense, Ranunculus acris), which suggests a significant increase of canopy openness caused by natural disturbances. There were also several alien species, which have invaded the plot during the last 25 years (e.g. Bidens frondosa, Conyza canadensis, Malus domestica). Some of the new species (e.g. Menyanthes trifoliata, Rhamnus cathartica, Rumex hydrolapathus) were recorded in the wettest places along Hwoźna river, therefore they could be not recorded by previous surveyors due to inaccessibility of the area, which was explored owing to the extremely dry vegetative seasons 2014 and 2015. Several species showed a tendency to expand their local ranges (increased number of inhabited plots): Deschampsia flexuosa, Euonymus europaeus, E. verrucosus, Frangula alnus, Malus sylvestris, Rubus nessensis, Rumex sanguineus, Sambucus racemosa, Ulmus glabra, Viscum album. Some other species, predominantly growing in pine forests, showed a tendency to decline, most probably due to serious stand disturbance caused by spruce bark beetle (Ips typographus) outbreaks: Calluna vulgaris, Chamaecytisus ruthenicus, Fragaria vesca, Genista tinctoria, Goodyera repens, Peucedanum oreoselinum, Scorzonera humilis, Vaccinium vitis-idaea.
Bryophytes. A small drop in the number of bryophyte species took place between the surveys (from 147 to 145). It was accompanied by low total species turnover (15), which suggests high stability of this taxonomic group. The most distinct difference between 1990 and 2015 is high number of records of Dicranum viride. The species is so distinctive that it must be the effect of recent colonization of the site by this rare species, which is protected by the Natura 2000 network (Wierzcholska et al. 2015). We showed that diverse, naturalistic forest assemblages buffer the effect of ash dieback on bryophyte communities – (Wierzcholska et al. in preparation). Ash-associated epiphytic bryophytes are able to exploit other niches in the diverse Białowieża Forest and thus far have not suffered a reduction in species richness at the temporal scale of our resurvey. The diversity of the Białowieża Forest generates resilience for these epiphyte populations, which themselves may be able to act as a source for the recovery of populations in managed forests and in order to support adaptation to the ongoing spread of ash dieback and to future threats.
Lichens. There were 159 species of lichen found in the limits of the study area in 1990. Of these, 151 species were confirmed in 2015 and 8 species, with low number of records in 1990, were not confirmed in the resurvey. This could be the result of having been overlooked in the field due to their rarity. In 2015 the total number of lichens was doubled: a total of 165 new species were recorded in the resurvey. Some of them were described as new to science during the past 30 years, others had not been recorded in Poland before 1990 (e.g.: Bacidia hemipolia f. pallida, Biatora pontica, Lecidella subviridis, Lecanora thysanophora, Protoparmelia hypotremella). Some new species records were the result of splitting some species recorded in the original survey (e.g.: Lepraria incanawas split into L. incana s.str., L. elobata, L. jackii, L. rigidula, L. eburnea, L. umbricola, L. ecorticata; Micarea prasinawas split into M. prasina s.str., M. byssacea, M. micrococca, M. hedlundii, M. soralifera, M. viridileprosa; Cetrelia cetrarioides into C. cetrarioides s.str., C. olivetroum, C. chicitae, C. monachorum; Cladonia chlorophaea into C. chlorophaea s.str., Cl. grayi, Cl. merochlorophaea, Cl. novochlorophaea and Graphis scripta into G. scripta s.str., G. betulina and G. pulverulenta). Some species were probably overlooked in the field in the original survey due to their small size or were mistaken for some other common species (e.g.: Fuscidea arboricola, Lecanora farinaria, Lecidella subviridis, Rinodina degeliana, R. efflorescens). The lichenologists working in the project (prof. Martin Kukwa and dr Anna Łubek) also found 10 species previously unknown to science, of which Micarea soralifera has been already described as new to science last year (Guzow-Krzemińska et al. 2016), while the description of the other species (Lecanora stanislai) is under preparation.
Due to the significant differences in the lichen taxonomy between the past and recent survey periods, all those species which were not known to the surveyors in 1990 were removed from all ecological analyses. It means that some species were grouped back into species sensu lato, e.g.:Lepraria incana s.l., Micarea prasina s.l., Cetrelia cetrarioides s.l., Cladonia chlorophaea s.l., Graphis scripta s.l., and some were not included (those described as new to science after 1990). After taxonomic correction, the number of lichen species for the entire study area still increased but only by 10%: from 140 (1990) to 156 (2015). The mean number of species per 1 ha plot increased significantlyfrom 39 ± 8 in 1990 to 61 ± 13 speciesin 2015.
General synthesis. The change in species composition and species richness differed between taxonomic groups and between forest communities. The vascular plants and lichens were characterized by high absolute species turnover (41 and 55 species respectively), expressing high dynamics in the studied ecosystems. There were plot-level extinctions, but they were not high. Bryophytes were the only group of species with neither extinctions nor colonisations registeredat the plot level, which is a sign of the stability of this taxonomic group in natural forest communities. However, the reaction of the studied taxonomic groups depended on the type of forest habitat: plots with net species extinctions were recorded only in the mixed deciduous forest of Tilio-Carpinetum community, which according to the literature, have a tendency to increase canopy cover. The total plot colonisations were at least twice as high as plot extinctions, with both characteristics highest for lichens. In effect the Bray-Curtis dissimilarity between the surveys was highest for lichens (0.44) and lowest for bryophytes (0.11).
We measured the reaction of the studied taxonomic groups to climate change and change in local environmental conditions by the analysis of species ecological indicator values (Zarzycki et al. (2002) for vascular plants, Ellenberg et al. (1992) for bryophytes and Wirth (2010) for lichens). There were significant indicator value increases in at least two of the three tested groups for light, temperature, soil reaction and nitrogen. Bryophyte temperature indicator increase and vascular plant reaction and nitrogen indicator increases were significantly higher in the pine forest of the Peucedano-Pinetum community in comparison with other communities. However, this significant effect was caused not by the climate change, which should influence all communities, but rather by a disturbance caused by spruce bark beetle outbreak, which killed the majority of spruces in the stand in 2002-2004 and allowed more light to reach the forest floor. Similar changes in lichen species diversity in wet types of forests: Carici elongatae-Alnetum and Circeao-Alnetumwere caused rather by the ash-dieback process than climate change. On the other hand the relationship between weighted mean indicator value change and phytosociological community at least for those taxonomic groups and indicators where analysis strongly implies a significant effect, showed that all communities are reacting in the same direction, just not all changes are significant in single communities (see Fig. 1). Therefore, it is probable, that climate effect is not very strong in the time scale of the quarter of a century, but becomes significant if reinforced by natural disturbances, such as insect outbreaks or fungal diseases causing large-scale tree diebacks. This hypothesis needs to be tested in a rigorous statistical manner. The synthesis paper (Jaroszewicz et al., in prep.) from resurveying of CRYPTO is well advanced and will be submitted in 2017 for publication (for details see Table E). The preliminary results have been presented at several conferences (see Table D3).
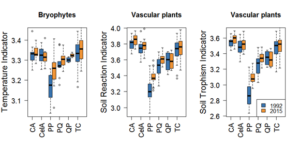
Figure 1. The samples of relationship between weighted mean indicator value change and phytosociological community (CA – Circaeo-Alnetum, CelA – Carici elongatae-Alnetum, PP – Peucedano-Pinetum, PQ – Pino- Quercetum, QP – Querco-Piceetum, TC – Tilio-Carpinetum) for indicators and taxonomic groups where analysis strongly implies a significant effect.